
![]()
![]()
![]()
Rỉ sắt
Giới thiệu chung
Lạc (đậu phộng) – Arachis hypogaea L. là một cây trồng quan trọng cho phần lớn cư dân trên trái đất này. Chính vì trong thành phần dinh dưỡng của nó chứa lipit (40 – 60%), protein (26 – 34%), gluxit (6 – 22%), chất xơ (2 – 4,5%) nên được sử dụng làm thực phẩm cho người, là nguồn nguyên liệu quan trọng cho công nghiệp chế biến dầu ăn thực vật.
 |
 |
 |
Trong hạt lạc có chứa tới 50% dầu không đông vì thế được sử dụng nhiều trong sản xuất bánh, kẹo, xà phòng. Sản phẩm sau khi chiết xuất dầu được coi là nguồn nguyên liệu quý cho sản xuất thức ăn gia súc. Thân lạc sau khi đã thu hoạch củ cũng là nguồn thức ăn cho gia súc có giá trị và bổ dưỡng. Lạc là một nguồn tuyệt vời của biotin (còn gọi là vitamin H hoặc vitamin B7), niacin (còn được gọi là vitamin B3, axit nicotinic hay vitamin PP), folate (vitamin B9), vitamin E, vitamin B1. Trong hạt lạc còn chứa can xi, sắt, magiê, kali, kẽm, phốt pho,….
Tuy nhiên, trên cây lạc có nhiều loại sâu, bệnh gây hại. Một trong những số đó là bệnh rỉ sắt. Hiện nay, ở những vùng trồng lạc trên thế giới rỉ sắt là một trong những tác nhân đem lại những thiệt hại lớn về kinh tế.
 |
 |
| Hình 1: Cánh đồng lạc không bị nhiễm bệnh (trái) và bị bệnh (phải) | |
Harrison (1972) lưu ý rằng tổn thất do rỉ sắt tại hai địa điểm ở Texas là 50% và 70%. Rỉ sắt gây thiệt hại nghiêm trọng cho cây lạc ở nhiều nơi trên thế giới với sản lượng củ giảm đến 70% có trong báo cáo của Harrison (1973); Subrahmanyam và McDonald (1987). Felix và Ricaud (1977) từ Mauritius báo cáo rằng thiệt hại do rỉ sắt lên tới 70% trong một số trường hợp. Zhou và cộng sự (1980) ghi nhận sản lượng giảm 20,4% cho lạc xuân và 17,3% cho lạc mùa thu ở tỉnh Quảng Đông của nước Cộng hòa Nhân dân Trung Hoa1. Trong nhiều địa phương của bang Maharashtra và Andhra Pradesh, Ấn Độ, rỉ sắt đã được báo cáo gây thiệt hại 40 –55% sản lượng củ đối với các giống mẫn cảm thường được trồng (Ghuge et al, 1981;. Mayee, 1987; Subrahmanyam và McDonald, 1987).2
Tác hại của bệnh này còn nghiêm trọng hơn khi nó xảy ra đồng thời cùng với bệnh đốm nâu do nấm Cercospora arachidicola Hori và bệnh đốm đen do nấm Cercosporidium persoratum (Berk. & Curt.) Deighton. Không có một loại thuốc trừ nấm nào có thể trừ cho cả rỉ sắt và hai bệnh đốm lá cùng một lúc. Do đó gián tiếp làm tăng chi phí sản xuất.
Ngoài việc giảm số lượng củ, giảm kích thước, trọng lượng củ thì bệnh hại còn thúc đẩy quá trình chín sớm từ 14 đến 28 ngày trong khi hạt lạc vẫn chưa chín đẫy (O'Brien, 1977).1,3 Bệnh còn làm giảm hàm hượng dầu trong hạt.1,3 Tổn thất về hàm lượng dầu do rỉ sắt đã được ước tính là khoảng 7% – 10% (Kenjale và cộng sự 1981).4 Trong một nghiên cứu tại Nigeria, hàm lượng protein đã được giảm 16 – 12% do các bệnh nấm trên lá bao gồm rỉ sắt (Salako và Adu, 1990).2
Tác nhân gây bệnh rỉ sắt lần đầu tiên được báo cáo từ Surinam năm 1827 (Leppik 1971). Nó lần đầu tiên được công nhận là Arachidis Uredo bởi Nils Gustav Lagerheim và sau đó có tên là Uromyces arachidis (Paul Hennings 1896). Năm 1922 Arthur và Mains đặt tên là Bullaria arachidis. Cuối cùng, các mẫu vật được thu thập bởi một nhà thực vật học người Pháp từ Caaguaza (Paraguay) đã được xác định bởi Spegazzini năm 1884 như một loài mới và đổi tên là Puccinia arachidis (Hennen et al. 1976).5
P. arachidis là tác nhân gây hại chuyên tính cao. Phạm vi ký chủ chỉ là lạc trồng (Arachis hypogaea) và một số cây dại thuộc họ Đậu như A. glabrata, A. burkartii, A. marginata, A. cardenasii, A. ehelods, A. stenosperma, A. nambyquarae và A. hagenbeckii.5,6
Từ trước 1969, bệnh rỉ sắt được giới hạn phía Nam Mỹ và Trung Mỹ. Tuy nhiên từ đó người ta thấy nó gây hại ở châu Á, Úc và Châu Đại Dương. Bệnh cũng đã được báo cáo từ năm 1974 từ một số nước châu Phi.1 Phân bố của bệnh theo địa lý như sau:
Châu Á: Bangladesh, Trung Quốc, Ấn Độ, Indonesia, Nhật Bản, Hàn Quốc, Cộng hòa Dân chủ Nhân dân Triều Tiên, Hàn Quốc, Malaysia, Myanmar, Nepal, Pakistan, Singapore, Sri Lanka, Philippines, Thái Lan, Việt Nam.
Châu Phi: Benin, Botswana, Burkina Faso, Côte d'Ivoire, Ethiopia, Ghana Guinea, Kenya, Malawi, Mali, Mauritius, Mozambique, Niger, Nigeria, Senegal, Somalia, Nam Phi, Sudan, Swaziland, Tanzania, Uganda, Zambia, Zimbabwe.
Trung Mỹ & Caribbean: Antigua và Barbuda, Belize, Cuba, Cộng hòa Dominica, Grenada, Guadeloupe, Guatemala, Honduras, Montserrat, Jamaica, Nicaragua, Panama, Puerto Rico, Saint Vincent, Grenadines.
Bắc Mỹ: Hoa Kỳ, Mexico.
Nam Mỹ: Argentina, Brazil, Colombia, Ecuador, Guyana, Paraguay, Peru, Suriname, Uruguay, Venezuela.
Châu Đại Dương: Úc, Queensland, Fiji, Papua New Guinea, Samoa, Quần đảo Solomon Tonga.
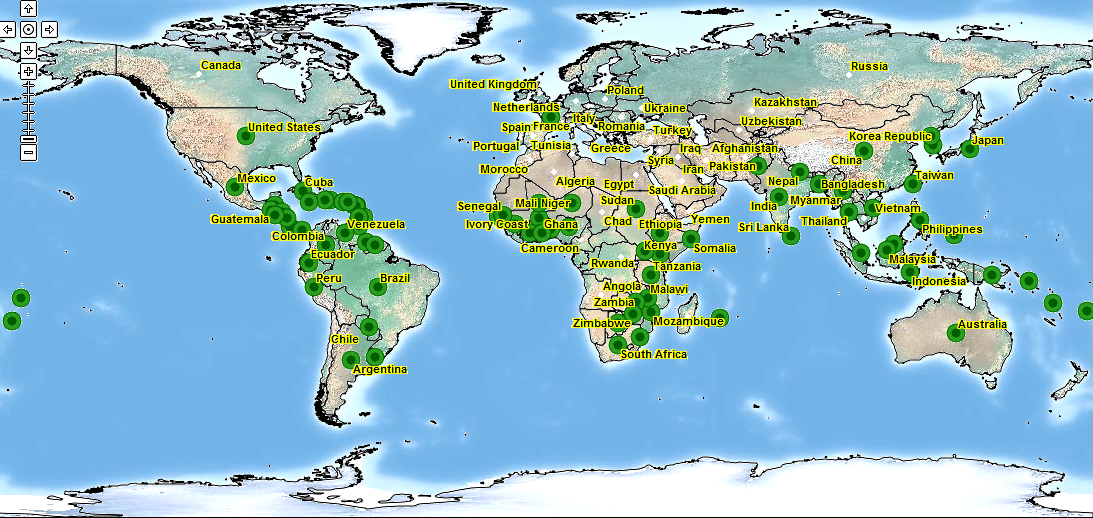
Hình 2: Phân bố địa lý gây hại của P. arachidis trên thế giới.
1.1 Triệu chứng
Sự xuất hiện của các triệu chứng của bệnh rỉ sắt lạc bắt đầu 8 – 10 ngày sau khi nhiễm với sự xuất hiện của các đốm trắng ở mặt trên của lá bánh tẻ. Một ngày sau đó, các đốm sáng màu vàng xuất hiện. Hình 3.
 |
 |
| Hình 3: Biểu hiện ban đầu của bệnh rỉ sắt lạc ở mặt trên của lá | |
Mặt dưới của các đốm màu đó xuất hiện các mụn mủ hơi nhô lên có kích thước như một điểm bút chì màu nâu sau chuyển sang màu cam đỏ. Hình 4.
 |
 |
| Hình 4: Các mụn rỉ sắt ở mặt dưới lá. | |
Các mụn mủ đó là các ổ (túi) bảo tử hạ – uredinia – thường tròn đến hình elip, đường kính thay đổi khác nhau trong khoảng 0,3 – 2,0 mm (hình 5) và thường được bao quanh bởi các mô lá có màu xanh nhạt đến nâu nhạt. Các mụn mủ mở rộng và vỡ sau khoảng 2 ngày kể từ ngày xuất hiện để lộ bào tử hạ – uredospores – hình tròn hoặc hình bầu dục, ban đầu màu cam tối sau trở thành màu nâu đậm.3,7
Hoại tử phát triển ở khu vực xung quanh các mụn mủ. Vùng hoại tử có thể hợp lại và có thể nhanh chóng gia tăng kích thước trong thời kỳ thời tiết ẩm ướt và nóng (hình 5).
 |
 |
 |
| Hình 5: Các vết bệnh phát triển. | ||
Bệnh phát triển làm lá bị khô, quăn và rủ xuống. Mặc dù các lá bị nặng nhanh chóng chuyển sang màu nâu và chết nhưng chúng vẫn gắn với cành cây.9,10
Cây bị nhiễm bệnh rỉ sắt nặng còi cọc, chuyển màu vàng. Cây bị nặng thường chết khô (hình 6, hình 1 – bên phải). Trên những cây đó củ lạc chín sớm và rụng, trọng lượng thấp.3,4,8

Hình 6: Ruộng lạc ở miền Nam Ấn Độ bị hại nặng bởi P. arachidis.
Ngoài lá, triệu chứng có thể phát triển trên tất cả các bộ phận trên mặt đất của cây (cuống lá, thân cây) và hoa, tia củ cũng không ngoại lệ.3,4,9,10
 |
 |
Nấm rỉ sắt thường xảy ra với nấm Cercopsora (đốm lá) và khi đó triệu chứng và tác hại của từng bệnh trở nên không rõ ràng.3 Tuy nhiên, nấm rỉ sắt phát triển với tốc độ nhanh hơn và khi bệnh nặng lá chết thường vẫn gắn ở trên cây, chứ không phải là làm rụng lá như đã thấy với Cercopsora.6
Những mụn mủ của bệnh rỉ sắt cũng dễ bị nhầm với tác hại của nhện.3,8
 |
 |
1.2 Nguyên nhân
Bệnh rỉ sắt lạc gây hại bởi nấm Puccinia arachidis Speg., Bộ Urediales, Lớp nấm đảm.
Trước khi mô tả các tính năng điển hình của bệnh rỉ sắt lạc điều cần thiết là cần nắm rõ chu trình sống chung của một Puccinia sp. điển hình. Một Puccinia sp. điển hình có tới 5 giai đoạn bào tử trong vòng đời. Bao gồm:
O – Đơn bào tử (Spermatia): sản sinh ra từ túi đơn bào tử (spermogonia).
I – Bào tử xuân (Aecidiospores): sản sinh ra từ túi bào tử xuân (aecidia).
II – Bào tử hạ (Urediniospores): sản sinh ra từ cụm bào tử hạ (uredinia).
III – Bào tử đông (Teliospores): sản sinh ra từ cụm bào tử đông (Telia).
IV – Bào tử đảm (Basidiospores): sản sinh ra từ đảm (basidium) do bào tử đông nảy mầm tạo thành.
Trong số năm giai đoạn này thì giai đoạn O và I (chỉ tồn tại trên ký chủ phụ) và giai đoạn IV (là giai đoạn sinh sản lưỡng bội) nhưng do P. arachidis là ký sinh chuyên tính và sinh sản vô tính nên nên chúng không tồn tại. Rỉ sắt lạc chủ yếu tồn tại trong tự nhiên ở giai đoạn II – uredial. Còn giai đoạn telial không phải là quá phổ biến và chỉ được báo cáo ở một số quốc gia ở Trung Mỹ, Florida và Brazil trong lá lạc hoang dã (Hennen et al. 1976). Nó cũng được quan sát gần đây từ lạc trồng tại Gadag, Karnataka, Ấn Độ (Tashildar et al. 2012).5,6
Trên đồng ruộng thông thường chỉ quan sát thấy các ổ bào tử hạ (uredinia) của P. arachidis (hình 7). Các ổ bào tử hạ nằm phân tán dưới biểu bì của cả hai mặt lá nhưng chủ yếu mặt dưới.8 Chúng nằm rải rác hoặc liên kết lại thành một nhóm. Bên ngoài được phủ bởi một lớp màng mỏng. Ban đầu màng mỏng đó bị rộp phồng lên. Sau đó vỡ ra tạo thành một lớp bột màu nâu đỏ đậm. Đó là các bào tử hạ. Trên những giống mẫn cảm, xung quanh những ổ bào tử hạ chính còn có những ổ bào tử hạ thứ cấp.6,7,9,10
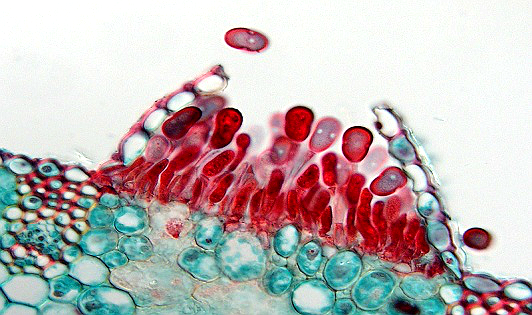
Hình 7: Ổ bào tử hạ.
Mỗi ổ bào tử chứa nhiều bào tử hạ màu nâu đỏ, hình tròn hoặc bầu dục và có cuống nhỏ: ngắn, mỏng và trong suốt. Hình 8,9.

Hình 8 (ảnh minh họa): Cuống và lỗ mầm của bào tử hạ nấm Puccinia nakanishikii.
 |
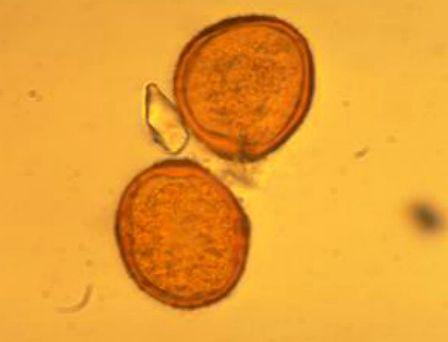 |
| Hình 9: Bào tử hạ của P. arachidis. | |
Vỏ ngoài bào tử hạ là vách mỏng (dày 1 – 2μm), có gai nhỏ, màu vàng nâu. Trong bào tử hạ thường có 2 – 4 lỗ mầm nằm trên đường xích đạo (giữa thân). Kích thước bào tử biến đổi khác nhau trong khoảng 18.56 – 33.00 x 17.47 – 26.48μm. Trung bình 24.96 x 21.22μm.4,7,11 Hình 10.
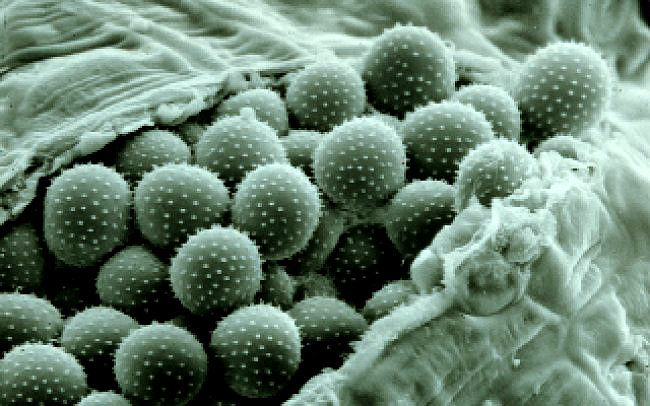 |
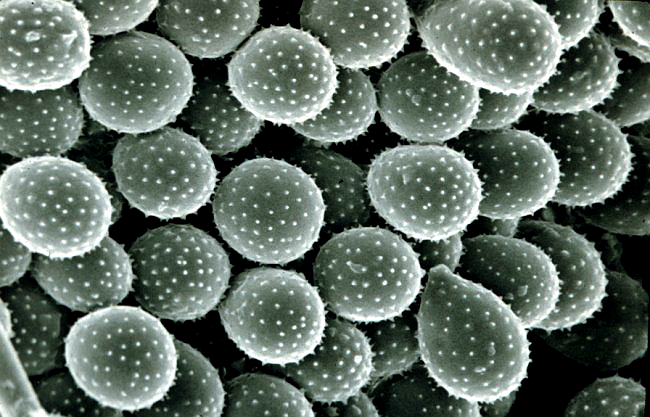 |
| Hình 10: Ổ bào tử hạ với các bào tử có gai nhỏ của Puccinia sp.. | |
Quá trình xâm nhiễm ban đầu của bào tử hạ P. arachidis ở mặt dưới của lá trên giống kháng – Arachis stenosperma V10309 – (a, b, c) và giống nhiễm – Arachis hypogaea cv. IAC-Tatu – (d, e, f, g) được thể hiện ở hình 11. Ảnh được chụp bằng kính hiển vi điện tử (Nguồn: Soraya Cristina De Macedo Leal-Bertioli, EMBRAPA, Brasilia, Brazil. © 2010 Blackwell Verlag GmbH).
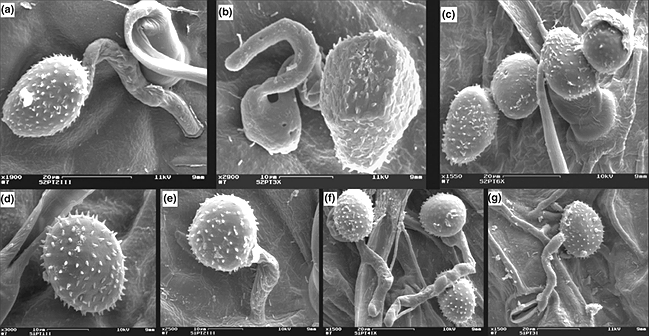
Hình 11: Quá trình xâm nhiễm ban đầu của P. arachidis
Chú thích:
d: Bào tử hạ trên lá của A. hypogaea cv. IAC-Tatu sau 3 giờ xâm nhiễm.
a, e: Ống mầm hình trụ, dài của bào tử hạ nảy mầm sau 6 giờ xâm nhiễm.
b, f: Giác bám (appressoria) hình thành sau 12 giờ xâm nhiễm.
c: Sự suy giảm tăng trưởng của ông mầm sau 24 giờ xâm nhiễm trên giống kháng.
g: Hiện diện của ống mầm bên lỗ khí khổng đang mở sau 72 giờ trên giống nhiễm.
Chu trình lây nhiễm của P. arachidis được thể hiện ở hình 12 dưới đây:
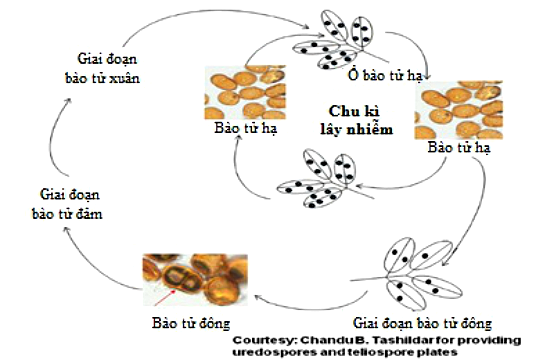
Hình 12: Chu trình lây nhiễm của P. arachidis.
Trên đồng, hạ bào tử lây nhiễm vào lá và hình thành các ổ bào tử hạ. Những ổ bào tử hạ sau khi “chín” vỡ và giải phóng nhiều hạ bào tử. Những hạ bào tử non trẻ lại bắt đầu một quá trình xâm nhiễm mới liên tục nhiều vòng liên tiếp trên lá lạc cho đến cuối vụ. Dưới nhiệt độ thấp và tình hình căng thẳng chất dinh dưỡng, các sợi nấm hai nhân (lưỡng hạch) của hạ bào tử thường hình thành cụm bào tử đông (Telia). Những Telia chứa nhiều bào tử đông (teliospores). Tuy nhiên, có rất ít báo cáo về sự tồn tại Teliospores của P. arachidis trong tự nhiên (Hennen và cộng sự 1976; Rodrigues và cộng sự 2006; Tashildar và cộng sự 2012).5,6
1.3 Phát sinh gây hại
Quá trình xâm nhiễm của hạ bào tử P. arachidis bắt đầu bằng việc một ống mầm (không phân nhánh) nảy mầm từ lỗ mầm nằm trên vách trong của màng bọc hạ bào tử. Ống mầm có đường kính khoảng 6µm, chiều dài khoảng 100 – 200µm (có thể dài đến 500µm). Ống mầm phát triển dọc theo đường rãnh bào (khoảng không giữa các vách tế bào). Sau khi tiếp xúc với một lỗ thoát, phần đầu của ống mầm hình thành một vòi bám (giác bám/ đĩa áp) hình elip, mỏng, có cùng kích thước với bào tử. (Vòi bám tích trữ một lượng lớn carbonhydrate – chủ yếu là glycerol – nên dễ dàng tạo ra một áp lực thẩm thấu cao giúp cho quá trình hút nước từ ngoài vào. Từ đó tạo ra một áp suất trương lớn, tạo điều kiện cho ống mầm phát triển vào trong các mô của tế bào. Ví dụ như vòi bám của nấm phấn trắng có thể tạo áp suất trương 20atm, của nấm đạo ôn khoảng 80atm, trong khi áp suất trương của lốp ô tô chỉ khoảng 2 – 3 atm).
Tiếp đó xảy ra một quá trình vận chuyển tế bào chất từ bào tử vào vòi bám đó. Vào cuối chu kỳ phát triển ở đầu vòi bám sưng lên. Thời gian từ ống mầm kéo dài đến hình thành giác bám thường được thực hiện trong vòng 12h đầu tiên của quá trình xâm nhiễm trên những giống mẫn cảm với bệnh. Từ vòi bám hình thành một móc (chốt) xâm nhiễm mảnh, hình elip giúp cho ống mầm xâm nhập vào khí khổng. Sau khi đi qua hết chiều dài của khí khổng móc xâm nhiễm tăng trưởng về kích thước hình thành một nang (túi) trong gian bào. Ở đây những sợi nấm hai nhân (lưỡng hạch) hình thành trong vòng 24h. Những sợi nấm đó hình thành các giác mút hình u tròn xâm nhiễm sang các tế bào liền kề. Quá trình xâm nhiễm hoàn thành sau 72h kể từ khi các ống mầm xâm nhập thành công vào bên trong tế bào. Quá trình xâm nhiễm được mô tả bằng hình 13 dưới đây:
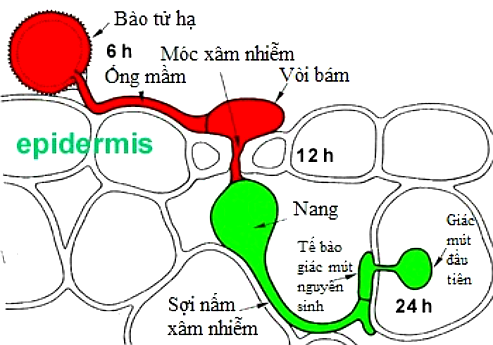
Hình 13: Quá trình xâm nhiễm của P. arachidis.
Sự tồn tại của P. arachidis trên tàn dư:
Hạ bào tử của P. arachidis tồn tại rất ngắn trên tàn dư cây lạc bị bệnh trong điều kiện nhiệt độ cao. Subrahmanyam và McDonald (1982) báo cáo rằng hạ bào tử tiếp xúc với thời tiết sau khi thu hoạch tại Trung tâm ICRISAT (The International Crops Research Institute for the Semi-Arid Tropics. Tạm dịch: Viện nghiên cứu nhiệt đới bán khô cằn quốc tế), chỉ sống sót 22 ngày sau khi thu hoạch trong vụ chính ở mùa mưa và chỉ 14 ngày sau khi thu hoạch vụ tăng cường. Mayee (1982) ghi nhận rằng trong điều kiện ở bang Parbhani (Ấn Độ), hạ bào tử mất khả năng tồn tại trong vòng 20 ngày. Mallaiah và Rao (1979b) báo cáo rằng trong các nghiên cứu của họ liên quan đến những mẫu bệnh sưu tập hàng tháng của các hạ bào tử từ các mảnh vụn bị nhiễm bệnh hơn một năm thì hạ bào tử không còn sức sống trong vòng bốn tuần.1,13
Hạ bào tử nếu không có ký chủ thì có một cuộc sống rất ngắn, ngoại trừ tại nhiệt độ rất thấp. Nó có thể được tồn tại ở nhiệt độ thấp mà không mất khả năng sống. Nhưng ở nhiệt độ 40oC thì khả năng sống sẽ nhanh chóng mất đi.4 Hạ bào tử nhanh chóng mất khả năng tồn tại ở nhiệt độ cao (28-35oC). Đây lại là phổ nhiệt độ phổ biến ở các vùng nhiệt đới (Subrahmanyam và McDonald, 1987).
Chính vì những nhận xét trên mà ta có thể nói rằng hạ bào tử không thể kéo dài thời gian sống trong vòng bốn tuần giữa các mùa vụ mà ở một số nước nhiệt đới và cận nhiệt đới, các mầm bệnh thực chất không tồn tại dưới hình thức hạ bào tử từ năm này sang năm khác.4
Do P. arachidis chỉ gây hại trên lạc và một số loài Arachis hoang dã nên việc canh tác liên tục là nguyên nhân tạo điều kiện cho bệnh phát sinh, phát triển quanh năm ở một số nước. Đặc biệt là ở Ấn Độ, Trung Quốc và Việt Nam.1
Sự lây truyền qua củ, hạt giống:
Kolte và Awasthi (1979) từ Uttar Pradesh, Ấn Độ, báo cáo rằng các cây con mọc từ những hạt giống của cây lạc nhiễm bệnh rỉ sắt nặng không thể hiện triệu chứng của bệnh trong điều kiện cách ly. Vilsoni (1980) từ Fiji, cũng cho rằng rỉ sắt không từ hạt gây ra. Subrahmanyam và McDonald (1982) trong thí nghiệm của mình đã chứng minh rằng rỉ sắt lạc không phải từ nội hạt gây ra.
Mặt khác, một số kết quả nghiên cứu cũng chỉ ra rằng rỉ sắt không rõ ràng chuyển từ mùa này sang mùa khác trên vỏ quả hay hạt giống và có rất ít cơ hội bệnh được lan truyền trên hạt giống. Đặc biệt là nếu các hạt giống được bảo quản ở nhiệt độ phòng bình thường trong 2 tháng (ICRISAT, 1978; Subrahmanyam và McDonald, 1982).
Tuy nhiên, trong thực tế hạt giống vẫn có thể bị nhiễm hạ bào tử của P. arachidis trên bề mặt qua con đường cơ học khi sử dụng củ thu hoạch từ vụ trước làm giống cho vụ liền kề. Do đó giải pháp xử lý hạt giống trước khi gieo trồng vẫn là cần thiết.
Khoảng cách lan truyền mầm bệnh:
Higgins (1956) báo cáo rằng nấm rỉ sắt dường như không qua mùa đông ở Mỹ, nhưng gió thổi từ vùng cận nhiệt đới là nguyên nhân cho bệnh phát sinh, phát triển. Wells (1962) cũng nhận thấy rằng rỉ sắt không qua mùa đông ở Bắc Carolina, (Mỹ) vì nó đã không tồn tại trên thân cây, ngọn cây sau khi thu hoạch, cây lạc chết hoặc trong đất. Ngoài ra, không có ký chủ khác của bệnh này đã được biết đến. Do đó, nguồn duy nhất của sự lây nhiễm từ các bào tử được mang đến từ các khu vực cận nhiệt đới nơi lạc được trồng vào mùa đông. Văn Arsdel và Harrison (1972) cũng đồng ý với nhận xét rằng hạ bào tử của P. arachidis được vận chuyển từ những nơi xa xôi đến Mỹ. Họ bắt bào tử trong nước mưa từ tháng Bảy đến tháng Tám (năm 1970) và quan sát bệnh rỉ sắt trong các khu vực gần nơi bẫy sau 10 – 15 ngày. Vào thời điểm đó, bệnh phát triển mạnh ở Mexico (cách 1290 km từ nơi quan sát của họ). Các tác giả nhận thấy rằng với với tốc độ gió 9 km/giây-1 trong 40 giờ đã có thể vận chuyển bào tử từ Mexico đến địa điểm đặt bẫy. Sau đó các bào tử sẽ được phân bố trên đồng ruộng bằng những trận mưa.8
Mayee và cộng sự (1977) ghi nhận tiến trình bệnh rỉ sắt phát sinh ở Ấn Độ như sau: bệnh xuất hiện trong tháng bảy – tháng tám ở miền Nam, trong tháng chín ở miền Trung và vào tháng mười ở miền Bắc. Subrahmanyam và McDonald (1982) dựa trên những quan sát của họ trên mô hình canh tác ở miền Nam Ấn Độ cũng đồng tình với nhận xét trên. Họ cho rằng lạc được trồng chỉ trong mùa mưa ở miền Bắc, nhưng lại được trồng quanh năm ở miền Nam. Do đó nguồn bệnh luôn luôn tồn tại. Dựa trên các mô hình trồng trọt ở Ấn Độ họ đưa ra giả thuyết rằng cây lạc trồng ở miền Nam có thể hoạt động như một nguồn của bệnh rỉ sắt. Từ đó bào tử được thực hiện bởi gió mùa để phân bố sang các vùng ở miền Bắc.
McDonald và cộng sự (1980) cho rằng ở Nigeria, rỉ sắt sống sót qua mùa khô ở miền Nam đã là nguồn lây lan trong mùa mưa đến các vùng trồng lạc chính của miền Bắc bằng gió. Ảnh hưởng của gió tây nam gây mưa liên quan đến sự sống còn của bào tử rỉ sắt và là nguyên nhân lan truyền chúng ở các quốc gia trồng lạc ở vùng Tây Phi.
Nói chung sự lây lan của bệnh rỉ sắt phụ thuộc vào gió để phát tán các bào tử và độ ẩm để tạo điều kiện cho nhiễm khuẩn ở bề mặt lá.
Các bào tử rỉ sắt cũng có thể lây lan nhờ con người và máy móc (Feakin, 1973).1
Ảnh hưởng của nhiệt độ đến quá trình nảy mầm hình thành bào tử và sự tồn tại:
Nhiệt độ là một trong những yếu tố có tác động nhất định đến quá trình nảy mầm của hạ bào tử.
Poudin và Macko (1974) từ Georgia, Mỹ, nhận xét rằng nhiệt độ tối ưu cho hạ bào tử nảy mầm là khoảng 18 – 20oC. Kono (1977) từ Nhật Bản tuyên bố rằng các hạ bào tử nảy mầm trên lạc lá tại 12 – 31oC với sự nảy mầm tối ưu ở 21 – 22oC, và quá trình phát triển thích hợp nhất tại 20 – 26oC. Pang (1977, 1982) từ Đài Loan tìm thấy rằng hạ bào tử nảy mầm ở 15 – 30oC với dải nhiệt độ tối ưu 20 – 25oC, và hạ bào tử được sản xuất 3 – 4 ngày sau khi hình thành của cụm bào tử hạ tại 15 – 30oC. Mallaiah và Rao (1979a) từ bang Andhra Pradesh, báo cáo rằng nhiệt độ tối ưu cho sự nảy mầm là khoảng 20oC. Zhou và cộng sự (1980) từ Cộng hòa Nhân dân Trung Hoa cho rằng nhiệt độ tối ưu cho bào tử nảy mầm là 24,5 – 28,0oC, tỷ lệ nảy mầm thấp ở nhiệt độ cao hơn 28oC và rất ít bào tử nảy mầm ở mức 11oC và 31oC. Không nảy mầm xảy ra ở dưới 8oC. Họ cũng nói thêm rằng các điểm chết nhiệt là 50oC trong 10 phút.1
Nhiệt độ trong khoảng 20-25oC là tối ưu cho hạ bào tử nảy mầm.7
Nhiệt độ cũng là một yếu tố quan trọng trong việc xác định tuổi thọ của hạ bào tử bệnh rỉ sắt. Nhiều thí nghiệm trong phòng nghiệm cho thấy hạ bào tử có thể được lưu trữ trong thời gian dài ở nhiệt độ thấp mà không mất khả năng tồn tại nhưng ở nhiệt độ cao thì ngược lại chúng nhanh chóng mất khả năng sống.
Veeranjaneyulu (1973) từ bang Andhra Pradesh, Ấn Độ, quan sát thấy rằng hạ bào tử có thể tồn tại trong chín tuần trên lá bị nhiễm bệnh được bảo quản ở nhiệt độ phòng (33 – 37oC), 13 tuần khi bị chôn vùi trong đất không cày ngay, 15 tuần ở 15oC và hơn 33 tuần ở 5oC. Mallaiah và Rao (1979b) thấy rằng hạ bào tử có thể tồn tại cho đến 4 tuần khi nhiệt độ đã xuống dưới 30oC, nhưng khi nhiệt độ trên 35oC thì bào tử không nảy mầm được ngay cả sau hai tuần. Zhou et al. (1980) đã nghiên cứu khả năng sống của hạ bào ở nhiệt độ khác nhau và trong các mùa khác nhau và nhận thấy rằng bào tử bị mất khả năng nhanh chóng khi ở nhiệt độ cao (40,5oC), 16 – 29 ngày với điều kiện nhiệt độ phòng mùa hè, 9 – 11 ngày ở 40oC, và 7 – 9 ngày ở 45oC. Tuy nhiên, các bào tử sống sót trong 120 – 150 ngày ở nhiệt độ mùa đông và mùa xuân, trong 3 – 6 tháng ở -24oC và trong gần một năm tại 5oC (Zhou và cộng sự, 1980). Subrahmanyam và McDonald (1982) nghiên cứu hạ bào tử được lưu trữ ở nhiệt độ -16, 6, 25, 40oC và ở nhiệt độ phòng có nhận xét rằng các bào tử giữ ở 40oC mất khả năng tồn tại trong vòng 5 ngày. Bào tử giữ ở 25oC sống sót cho đến 40 ngày và những bào tử được lưu trữ ở nhiệt độ phòng (hạt giống) sống sót trong 45 ngày. Bào tử được lưu trữ tại 6oC có thể sống sót 60 ngày còn tại -16oC thì khả năng sống hơn 120 ngày.1
Quá trình lây nhiễm cũng chịu tác động của nhiệt độ. Quá trình này của hạ bào tử diễn ra thành công tại 22oC – 26oC. Khoảng 8 –10 ngày sau khi quá trình ủ bệnh xảy ra thì các triệu chứng trở nên rõ ràng trên bề mặt lá. Lúc này một chu trình gây bệnh mới bắt đầu.4 Mallaiah và Rao (1979a) cho thấy rằng sự phát triển rỉ sắt xảy ra trong một phạm vi rộng của nhiệt độ 20 – 35oC, thích hợp nhất ở 25oC. Thời gian ủ bệnh là chỉ 7 ngày ở 25oC, trong khi đó là 14 ngày ở 35oC. Munde và Mayee (1979) cho rằng thời kỳ ủ bệnh được kéo dài ở nhiệt độ cao. Họ phát hiện ra rằng ở 23oC, thời gian ủ bệnh thay đổi từ 6 – 9 ngày. Ở nhiệt độ 27oC nó tăng lên đến 8 – 10 ngày và ở 30oC nó là 11 – 14 ngày. Zhou và cộng sự (1980) đã chỉ ra rằng thời gian ủ bệnh của bệnh rỉ sắt thay đổi dưới nhiệt độ khác nhau. Nó có thời gian ủ bệnh 18 ngày tại 18oC, 10 – 14 ngày ở 24oC, 6 – 8 ngày tại 24,5 – 26oC và 9 ngày ở 29oC.
Bệnh tiến triển chậm ở 10oC và 35oC do thời kỳ ủ bệnh bị kéo dài (Mallaiah và Rao 1979; Rao và cộng sự 1997; Zhou 1987).7
Ảnh hưởng của nhiệt độ và ẩm độ đến sự phát triển của bệnh:
Castellani (1959) tại Cộng hòa Dominican lây nhiễm nhân tạo bằng hạ bào tử rỉ sắt, bảo quản bốn ngày trong phòng thí nghiệm với độ ẩm tương đối 80 – 90% và nhiệt độ 28 – 32oC. Cây được sau đó được trồng trong điều kiện tự nhiên (nhiệt độ tối thiểu 26oC). Các triệu chứng xuất hiện 12 – 14 ngày sau khi cấy. Bromfield và Cevario (1970) tại Hoa Kỳ nhận xét rằng phạm vi nhiệt độ từ 25 – 30oC và độ ẩm tương đối trên 80% thuận lợi cho rỉ sắt phát sinh.1
Mallaiah và Rao (1979a) cho rằng hạ bào tử không nảy mầm khi độ ẩm tương đối là dưới 100%. Tuy nhiên, đối với bào tử nảy mầm trên bề mặt lá, sự hiện diện của nước tự do là tối cần thiết. Các hạ bào tử không nảy mầm ở một độ ẩm dưới 100%. Nếu độ ẩm tương đối trên 80% thì hạ bào tử chỉ nảy mầm khi được đặt trong một màng mỏng của nước (Cook 1980b).1,4
Munde và Mayee (1980) ở Maharashtra (Ấn Độ) thấy rằng một khi xâm nhiễm đã xảy ra, bệnh tiếp tục phát triển tốt ở nhiệt độ 30oC và độ ẩm tương đối 80%. Zhou và cộng sự (1980) giữ cây cấy trong tủ ẩm cho 4, 6, 8 và 24 giờ ở 25,5 – 26,0oC xâm nhiễm xảy ra trong vòng 4 giờ, nhưng mức độ nghiêm trọng còn thấp. Khi cây được sống trong các phòng ẩm với thời gian hơn 6 giờ, bệnh đạt 100%. Họ cũng báo cáo rằng hạ bào tử chỉ nảy mầm khi tiếp xúc với những giọt nước và nếu không thì không thể nảy mầm ngay cả khi trong điều kiện độ ẩm bão hòa.1
Mallaiah và Rao (1982) từ bang Andhra Pradesh (Ấn Độ), thấy rằng rỉ sắt phát triển nhanh khi nhiệt độ là từ 28 đến 34oC và độ ẩm tương đối từ 55 đến 85%. Subrahmanyam và cộng sự (1984) cho biết nhiệt độ trong khoảng 20 – 30oC, nước tự do trên bề mặt lá và độ ẩm tương đối cao là thuận lợi cho quá trình lây nhiễm và phát triển của bệnh tiếp theo. Độ ẩm tương đối > 80% được hỗ trợ nảy mầm cao hơn khi bào tử được đặt trên một màng mỏng của nước trên bề mặt lá.1,7,13
Mưa là yếu tố tạo nên độ ẩm không khí tương đối và tạo nên màng nước trên bề mặt lá. Trong điều kiện lượng mưa và độ ẩm của vụ lạc cuối ở Honduras và Nicaragua (Trung Mỹ), bệnh trở nên nghiêm trọng và sau đó nó trở nên khó kiểm soát (Arneson 1970). Ở Venezuela, rỉ sắt trở nên nghiêm trọng khi cây lạc được trồng trong mùa mưa hoặc khi sương dồi dào (Hammons 1977). Tại Ấn Độ, một khoảng thời gian khô liên tục đặc trưng bởi nhiệt độ cao (> 26oC) và độ ẩm tương đối thấp (<70%) được báo cáo là nguyên nhân làm chậm quá trình phát triển của rỉ sắt. Nhưng mức độ trở nên nghiêm trọng hơn khi mưa liên tục, độ ẩm tương đối cao và nhiệt độ 20 – 26oC (Siddaramaiah và cộng sự 1980).4
Krishna Prasad et al. (1979) từ Dharwad (Ấn Độ) thấy rằng bệnh sẽ phát sinh thuận lợi nếu nhiệt độ tối ưu là 23 – 24oC, với mưa liên tục dẫn đến độ ẩm tương đối trung bình > 87% trong một vài ngày. Dưới những điều kiện này, quá trình xâm nhiễm xảy ra rất nhanh và các triệu chứng ban đầu được ghi nhận trong vòng 10 – 12 ngày. Siddaramaiah và cộng sự (1980) lưu ý rằng thời kỳ khô hạn liên tục với nhiệt độ trên 26oC và độ ẩm dưới 70% thì mức độ nghiêm trọng của bệnh sẽ bị trì hoãn.1
Người ta cũng đã được chứng minh rằng mưa nhẹ phát tán bệnh hơn mưa rào vì số lượng bào tử trên các vết bệnh trong tán cây đã giảm mạnh sau những cơn mưa rào (Savary và Janeau 1986).7
Ngoài ra còn một số điều kiện khác ảnh hưởng đến quá trình phát sinh, phát triển và gây hại của bệnh.
Tính mẫn cảm của cây lạc còn liên quan đến tuổi cây. Theo McVey (1965), với các nhà máy trở nên dễ bị bệnh nhất ở 5 – 6 tuần tuổi . Mặc dù bệnh có thể lây nhiễm ở tất cả các tuổi cây.3,4
Ánh sáng đã được xác nhận có ảnh hưởng ức chế sự nảy mầm hạ bào tử và kéo dài ống mầm ống. Các thí nghiệm lây nhiễm nhân tạo chỉ ra rằng có thể sẽ thành công hơn nếu thực hiện vào buổi tối chứ không phải vào ban ngày.13 Mallaiah (1976) ghi nhận rằng cây được trồng trong bóng râm phát triển mụn mủ rỉ 1 – 2 ngày sớm hơn so với cây trồng dưới ánh sáng mặt trời trực tiếp. Ông cũng ghi nhận rằng thời gian ủ bệnh trong những tháng mùa hè (tháng năm và tháng sáu) là 18 ngày, trong khi trong những tháng mùa đông (tháng mười hai và tháng giêng) đó là chỉ 7 ngày. Munde và Mayee (1980) nhận thấy rằng cây lạc khi được lây nhiễm và trong điều kiện ở nhiệt độ 27oC và độ ẩm tương đối 100% trong 120 giờ, theo một chu kỳ của ánh sáng huỳnh quang 12 giờ sáng và 12 giờ tối thì khả năng lây nhiễm rỉ đạt tối đa với sự xuất hiện sớm của mụn mủ. Đối với bệnh rỉ sắt lạc quá trình lây nhiễm của một hạ bào tử từ lúc nảy mầm cho đến khi xâm nhiễm có thể mất 16 – 20h, dưới điều kiện nhiệt độ tối ưu là 22 –25oC, ánh sáng thấp và độ ẩm trên bề mặt lá kéo dài.10
Mật độ hạ bào tử tập trung cũng ảnh hưởng đến sự nảy mầm và những giai đoạn tiếp theo trong quá trình gây nhiễm.4
Khả năng tồn tại của hạ bào tử trên tàn dư còn phụ thuộc vào đặc tính của giống nhiễm hay giống chống chịu, kháng. Còn phụ thuộc vào thời vụ và điều kiện bảo tồn của chúng.1
Ở miền Bắc Việt Nam bệnh phát sinh gây hại quanh năm nhưng thường gây hại nặng hơn trong vụ lạc thu, thu đông so với ở vụ xuân.
Để quản lý dịch hại cần áp dụng các biện pháp:
Biện pháp canh tác
Thu thập các tàn dư cây sau khi thu hoạch và tiêu hủy bằng cách đốt.
Mặc dù rỉ sắt có tính chuyên hóa cao nhưng cũng cần lưu tâm đến những cây ký chủ phụ của bệnh này trước và sau khi thu hoạch để ngăn chặn nguồn bệnh.
Trong mỗi địa phương không nên trồng liên tiếp hai vụ lạc trên cùng một cánh đồng. Khoảng thời gian giữa hai vụ cũng phải bảo đảm cách nhau từ 1 tháng trở lên. Xuống giống tập trung.
Luân canh có thể phá vỡ chu kỳ bệnh bởi vì rỉ sắt lạc là nấm bệnh chuyên tính. Nếu có thể ở mỗi địa phương điều chỉnh công thức luân canh chung sao cho lạc không phải trồng vào thời điểm mưa nhiều, nhiệt độ ôn hòa (20 – 25oC).
Sử dụng củ giống khỏe mạnh. Xử lý hạt giống trước khi gieo.
Bệnh rỉ sắt có tương quan với độ ẩm tương đối cao nên tránh tưới nước trên cao vào chiều mát, giảm mật độ cây trồng, làm sạch cỏ dại (vì cỏ dại tăng trưởng mạnh sẽ tạo độ ẩm cao trong tán cây).
Trên những ruộng trồng lạc (nhất là những vùng chuyên canh) thì cần tăng cường vôi và phân lân (60 – 75kg lân/ ha) .
Kiểm tra đồng ruộng thường xuyên, ít nhất hai lần một tuần. Quan sát, theo dõi những chiếc lá đang chuyển màu vàng trước thời gian của chúng. Phát hiện kịp thời những đốm màu đỏ cam ở mặt dưới của lá.
Để dự báo khả năng xuất hiện gây hại và bùng phát thành dịch có thể dựa vào một số chỉ tiêu sau: nhiệt độ trung bình vào khoảng 20 – 22oC, độ ẩm tương đối > 85%, mưa ba ngày mưa trong một tuần và liên tiếp trong 2 tuần (Mayee, 1983).
Cần thực hiện kiểm dịch thực vật (đối nội, đối ngoài) nghiêm ngặt để tránh sự lây lan của các bào tử trên vỏ quả hay hạt (xâm nhiễm một cách cơ học).
Cuối cùng, một biện pháp rất hữu hiệu là sử dụng giống kháng. Giống kháng là một chiến lược quan trọng trong việc giảm tổn thất năng suất do rỉ sét. Kháng rỉ sắt được đặc trưng bởi tăng thời gian ủ bệnh, làmgiảm tần suất nhiễm trùng, giảm đường kính tổn thương, và chỉ số hình thành bào tử thấp. Trên các giống kháng các ống mầm chết mà không thể phát triển hơn nữa. Sự khác biệt của giống kháng còn được thểhiện bởi sự khác biệt về tốc độ và mức độ phát triển của các sợi nấm trong các mô lá.
Biện pháp thuốc BVTV
Sử dụng thuốc BVTV theo 4 đúng.
(Liên hệ với chúng tôi)
D.A.M